Activités de Recherche
Le sujet de ma thèse a été l'effet des oxystérols sur les astrocytes réactifs et tumoraux.
Lors d'une lésion du système nerveux central (SNC) entraînant une mort neuronale par suite d'un traumatisme, d'une maladie neurodégénérative ou d'une lésion neurotoxique, les cellules astrogliales (non-neuronales) deviennent hypertrophiques et hyperplasiques. Les astrocytes changent de morphologie, elles deviennent fibreuses avec de nombreux prolongements ramifiés et sont riches en Glial Fibrillary Astrocytic Protein (GFAP), le principal filament intermédiaire astrocytaire. Une sous-population d'astrocytes entame une prolifération mitotique. Les astrocytes morphologiquement transformés et/ou mitotiques sont appelés astrocytes réactifs et participent à la gliose réactionnelle. Ces cellules forment ainsi une cicatrice gliale qui délimite le site de la lésion du SNC, ce tissu dense formé d'astrocytes réactifs peut contribuer à empêcher la régénération des neurones en bloquant la repousse axonale. Dans le cadre de ma thèse j'ai étudié l'effet des oxystérols sur les astrocytes réactifs in vivo et in vitro.
Les oxystérols sont des dérivés oxygénés du cholestérol, ils portent une fonction hydroxyl ou cétone supplémentaire. Le 7ß-hydroxycholestérol (7ßOHCH) est cytotoxique envers des astrocytes tumoraux en culture sans être toxique pour des astrocytes normaux ou des neurones en culture primaire. L'effet toxique est relié à la synthèse par la cellule d'un métabolite où la fonction hydroxyl en C-3 est estérifiée par un acide gras (Behr et al. , 1991, FEBS Letters 280 : 202-206).
J'ai démontré in vitro que le 7ßOHCH et certains 7ßOHCH-3-esters réduisent la gliose réactionnelle induite par une lésion dans le cortex frontal de rats âgés de 6 jours et adultes (Bochelen et al. , 1992, Neuroscience 51 : 827-834). La molécule la plus efficace, le 7ßOHCH-3-oleyl inhibe à la fois la transformation morphologique des astrocytes (Gimenez y Ribotta M. et al. , 1995, J. Neurosci. Res. 41 : 79-95) ainsi que leur prolifération (Bochelen et al. , 1992, Neuroscience 51 : 827-834). Les études de relation structure-activité ont montré une relation entre le potentiel d'inhibition de la biosynthèse du cholestérol in vivo par les différents analogues structuraux et leur efficacité dans l'inhibition de la gliose et de la prolifération des astrocytes (Bochelen et al. , 1995, J. Neurochem. 65 : 2194-2200).
L'injection locale de 7ßOHCH-3-oleyl inclus dans des liposomes après hemisection de la moelle épinière favorise la repousse dans la moelle lésée de fibres sérotoninergiques (Gimenez y Ribotta M. et al. , 1995).
Lors du stage post-doctoral au sein du Laboratoire de Neurobiologie Moléculaire des Interactions Cellulaires au Centre de Neurochimie à Strasbourg j'ai étudié in vitro l'effet des oxystérols sur les astrocytes réactifs.
Pour cela j'ai utilisé un modèle in vitro d'astrocytes réactifs. Le traitement d'astrocytes en culture primaire par du dibutyryl-AMP cyclique (db-AMPc) ou un agoniste des récepteurs b-adrénergiques, induit une élévation du taux intracellulaire d'AMPc et une transformation morphologique des cellules qui deviennent analogues aux astrocytes réactifs. Le 7ßOHCH est toxique envers ces astrocytes morphologiquement transformés sans affecter des astrocytes normaux. L'inhibition de l'augmentation de l'AMPc par l'antagoniste des récepteurs ß-adrénergiques abolit la cytoxicité du 7ßOHCH. Le dosage de l'activité de la protéine kinase dépendante de l'AMPc (PKA) a démontré que la PKA était inhibée dans les minutes suivant le double traitement par le db-AMPc et le 7ßOHCH. Le facteur de transcription CREB est un des substrats de la PKA, l'inhibition de cette kinase peut donc affecter l'expression de certains gènes comme celui de la NCAM ou de la GFAP.
Au sein de l'unité Neurodégénération de la recherche préclinique de Novartis Pharma à Bâle je me suis consacré à l'étude de l'ischémie cérébrale.
J'ai caractérisé un modèle expérimental d'ischémie cérébrale réversible chez le rat et la souris. Cette technique consiste à bloquer le flux sanguin dans l'artère cérébrale moyenne (MCA) au moyen d'une suture en nylon introduite dans la partie extracrânienne de l'artère cérébrale interne. Par cette technique la boîte crânienne n'est pas endommagée limitant l'impact pour l'animal de l'opération et permet une reperfusion rapide et bien controlée de l'artère.
J'ai caractérisé le modèle dans une souche de rats hypertendus (SHR). Nous avons mesuré l'évolution du flux sanguin cérébral à l'aide d'un traceur radioactif et de méthodes non-invasives d'Imagerie par Résonance Magnétique Nucléaire (Allegrini P., et al. Quantitative measurement of regional cerebral blood flow: Correlation of MRI perfusion measurements using arterial spin tagging or bolus tracking with the 14C-iodoantipyrine method; 6th meeting of the International Society for Magnetic Resonance in Medecine, April 1998, Sydney). Les conséquences sur le cerveau de périodes ischémiques de durée variable (60, 120, 180mn ou permanent) ont été mesurées à des temps précoces (24-48h) à l'aide de l'IRM ; ces résultats ont été corrélés avec les résultats obtenus post-mortem dans les mêmes animaux par coloration histologique à des temps plus tardifs (1-3 semaines).
J'ai utilisé ce modèle pour tester des substances potentiellement neuroprotectives tels les immunosuppresseurs FK506, ASM-981, rapamycine et cyclosporine A. J'ai ainsi démontré que l'effet neuroprotecteur du FK506 et son dérivé ASM 981 sont lié à l'inhibition de la phosphatase calcineurine (Bochelen et al., 1999, J. Pharmacol. Exp. Therap. 288 : 653-659).
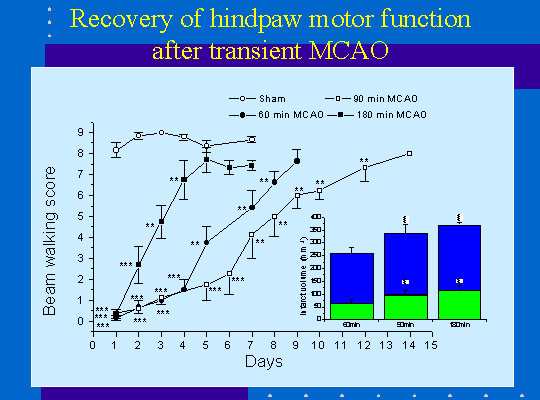
J'ai étudié les conséquences neurologiques, essentiellement d'ordre sensori-moteur, de la lésion ischémique à l'aide des tests du "beam-walking" et du "staircase"(figure1).
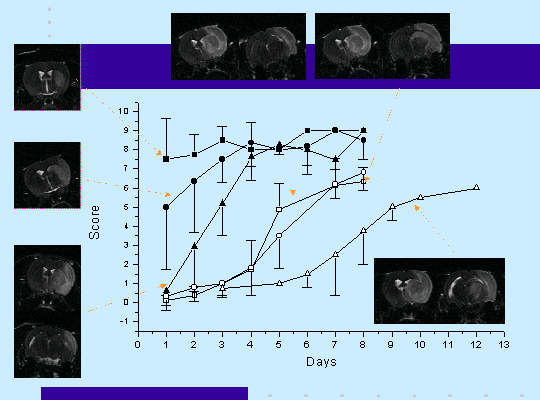
L'occlusion transitoire de la MCA produit des déficits moteurs important au niveau de la patte postérieure controlatérale à la lésion ainsi que le montre le test du beam-walking (figure 2) . Ce déficit fonctionnel s'améliore au cours du temps et les animaux reviennent à un niveau de performance proche de la normale. Le temps nécessaire à la récupération fonctionnelle dépend de la durée de l'occlusion de la MCA et par conséquent du volume de la lésion (figure 2) particulièrement dans le cortex somatosensoriel . Une lésion incluant l'hippocampe n'a pas de conséquence sur la récupération fonctionnelle par contre lorsque des structures thalamiques sont touchées celle-ci est retardée voire empêchée (figure 3).
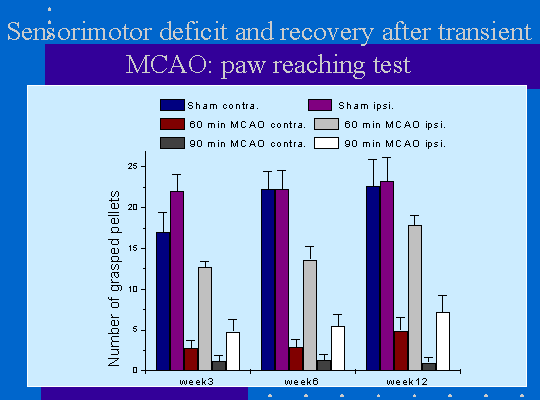
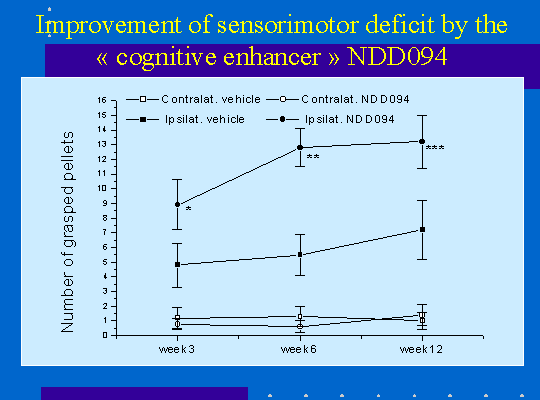
Les lésions ischémiques induisent également un déficit neurologique des capacités de préhension des pattes antérieures mesurée par le test du staircase. Le déficit de la patte antérieur controlatérale est très sévère et irréversible pendant la période considérée (15 semaines, figure 4). La patte ipsilatérale à la lésion présente également un déficit mais moins marqué. Dans le groupe d'animaux ayant subi 60mn d'ischémie une récupération fonctionnelle est observée au cours du temps (figure 4). Cette amélioration du déficit peut être facilitée par un traitement pharmacologique (figure 5).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}